欧洲玉米螟 (Ostrinia nubilalis) 幼虫肠道表达序列标签:探究候选基因与苏云金芽孢杆菌毒素和抗性潜在的关联 
 作者
作者  通讯作者
通讯作者
基因组学与应用生物学, 2010 年, 第 29 卷, 第 4 篇 doi: 10.5376/gab.cn.2010.29.0004
收稿日期: 2010年07月28日 接受日期: 2010年08月15日 发表日期: 2010年12月06日
本文首次发表在BMC Genomics开放取阅期刊上。本文是依据《Creative Commons Attribution License》协议,用中文再次发表与传播,如果读者对中文含义有歧义的话,请以英文原文为准。如果任何人要引用本研究内容,建议的引用格式如下:
Chitvan et al 2009, Expressed sequence tags from larval gut of the European corn borer (Ostrinia nubilalis): Exploring candidate genes potentially involved in Bacillus thuringiensis toxicity and resistance,BMC Genomics 2009, 10: 286 (doi: 10.1186/1471-2164-10-286)
基因组学与应用生物学,2010年,第29卷,第4篇 doi: 10.5376/gab.cn.2010.29.0004
背景
目前,鳞翅目类昆虫已经超过16 000种,其中包括一些对农作物、森林以及储存物最具破坏性的害虫。然而,我们对鳞翅目昆虫基因组信息的了解却十分有限,只有极少数的科学研究将重点放在构建鳞翅目昆虫幼虫肠道表达序列标签文库上。对昆虫肠道的基因表达的认识是了解食物消化基本生理学的关键,通过探索它们与苏云金芽孢杆菌相互作用的类型、方式等,可以找到新毒素的新靶标进而用于控制害虫。欧洲玉米螟(ECB, Ostrinia nubilalis)是北美和西半球作物生产中对玉米危害最大的害虫之一,本研究所分析的表达序列标签来自于ECB的幼虫肠道中。我们的研究目的是建立一个欧洲玉米螟幼虫肠道的特异性表达序列标签数据库,作为未来研究的基因组资源,同时从欧洲玉米螟中发掘与Bt相互作用和对Bt产生抗性的潜在的相关联的候选基因。
结果
我们用欧洲玉米螟5龄幼虫的肠道作为材料,构建了两个cDNA文库,并从这两数据库中通过测序,总共获得了15 000个EST序列。在获得的EST序列中,共有12 519 (83.4%)个ESTs为高质量片段,平均长度为656 bp。这些ESTs象征着有2 895个特异性序列,这些特异性列中包括1 738个单体和1 157个重叠群。在这些特异性序列中,62.7%编码假定蛋白质,这些假定蛋白的序列与Genbank数据库已有可用于比对的序列有重要的相似性(E-value ≤ 10<sup>-3</sup>)。我们对EST分析表明有52个候选基因可能在Bt毒性和抗性中的作用中扮演角色。这些基因编码了18种胰蛋白酶,18种糜蛋白酶,13种氨基肽酶,2种碱性磷酸酶以及一种钙粘着蛋白。我们利用RT-PCR的方法比较对Cry1Ab敏感系和具抗性虫系的欧洲玉米螟中的41个候选基因的表达谱,实验结果表明,与敏感虫系相比,抗性虫系中的2种胰蛋白酶基因和2种糜蛋白基因和一种氨基肽酶基因表达量下降明显。相反,在抗系虫系中3个胰蛋白酶基因,3个糜蛋白酶基因,2个氨肽酶基因以及2个碱性磷酸酶基因的的表达量增加。候选基因如此大的表达差异可能说明它们与Cry1Ab抗性之间存在关联。实际上,在先前研究中已经发现了一些胰蛋白酶和糜蛋白酶可以激活或抑制Bt的原毒素和毒素。然而,在其它一些昆虫种类体内,一些氨肽酶,钙粘着蛋白以及碱性磷酸酶已经被证实是作为Bt的毒素蛋白受体。
结论
我们利用从欧洲玉米螟幼虫的肠道中获得的15 000个cDNA构建了一个相对比较大的EST数据库,该数据库由12 519个高质量的序列组成。据我们了解,该数据库是鳞翅目害虫中最大的肠道特异性EST数据库。我们的研究也为以后欧洲玉米螟肠道特异性DNA基因芯片的开发、利用等研究奠定基础,而这种基因芯片可以用来分析Bt原毒素/毒素的基因表达的整体变换以及Bt抗性虫系和感性虫系之间的遗传差异。此外,我们鉴定了52个可能与Bt的毒性和抗性有关的候选基因。通过RT-PCR检测,我们从获得的41个候选基因选出15个有表达差异的基因,其中有5个基因在Cry1Ab抗性虫系中是表达量减少,而其余10个基因是表达量增加。这些结果可能会使我们坚信这些候选基因与Bt抗性有关,并为探讨cry1Ab在欧洲玉米螟中的抗性机制提供了新的观点。
在最近的几年里,昆虫基因组信息日益增加。一些昆虫物种的全基因组已经被测序,包括果蝇(Drosophila melanogaste) (Adams et al., 2000)、按蚊(Anopheles gambiae) (Holt et al., 2002)、埃及伊蚊(Aedes aegypti) (Nene et al., 2007)、意蜂(Apis mellifera) (4)、蚕(Bombyx mori) (Mita et al., 2004; Xia et al., 2004)、红粉甲虫(Tribolium castaneum) (7)以及11种其他的果蝇属昆(Crosby et al., 2007; Lin et al., 2007)。其他昆虫如豌豆蚜虫(Acyrthosiphon pisum),尖音库蚊(Culex pipiens),三种寄生蜂(Nasoniasp),黑森瘿蚊(Mayetiola destructor),吸血虫(Rhodnius prolixus),以及体虱(Pediculus humanus) (Deng et al., 2006; Grimmelikhuijzen et al., 2007; Sattelle et al., 2007)等物种的全基因组测序目前正在进行中。红粉甲虫是目前为止进行全基因组测序的昆虫物种中唯一农业害虫。
鳞翅目昆虫的种类数仅次于鞘翅目昆虫,是第二大昆虫群体,它有超过16 000种昆虫,其中包含了对农作物、森林和储藏物最具破坏性的害虫(Pierce, 1995)。家蚕是第一个完成全基因组测序的鳞翅目昆虫(Xia et al., 2004) 。但是其他鳞翅目昆虫的基因组信息还很有限,尤其是一些农业害虫的基因组信息,而这些信息由于它们经济效益的重要性和生物多样性而迫切需求。ESTs测序被认为是鉴定大量的表达基因的一种非常经济实用的方法,它可以用于基因表达和其他基因组研究上(Gerhold and Caskey, 1996; Dimopoulos et al., 2000; Porcel et al., 2000)。其实,ESTs已经可以从一些鳞翅目昆虫内获得,如:家蚕(Mita et al., 2003)、云杉色卷蛾(Choristoneura fumiferana) ( Li et al., 2003, http://www.pestgenomics.org/database.htm)、棉铃虫(Helicoverpa armigera)、 (Dong et al., 2007)、小菜蛾(Plutella xylostella) (Eum et al., http://www.uniprot.org/uniprot/Q60FS0) 、烟草天蛾(Manducasexta) (Robertson et al., 1999; Zou et al., 2008)和草地粘虫(Spodoptera frugiperda) (Deng et al., 2006; Nègre et al., 2006)。
我们在寻找新的昆虫害虫防治方法上,人们早就认识到在寻找新昆虫害虫防治策略时,昆虫肠道是一种很重要的研究靶标。然而直到现在,仅仅少数科学研究致力于将鳞翅目昆虫肠道特异性EST数据库作为一种工具来认识候选基因与杀虫剂的毒性和杀虫剂抗性的有关的研究。苹浅褐卷叶蛾(Epiphyas postvittana) (6416 ESTs) (Simpson et al., 2007),披肩粘虫(Mamestra configurata) (30种与丝氨酸蛋白酶相关的序列) (Hegedus et al., 2005),以及玉米螟(欧洲玉米螟, Ostrinia nubilalis) (1745 ESTs) (Coates et al., 2008)的肠道特异性EST文库已经被报道过。
欧洲玉米螟是危害最为严重玉米害虫之一,每年仅在美国就会造成10亿美元的经济损失(Ostlie et al., 1997; Gould, 1998)。欧洲玉米螟代表一个秸秆食心虫群,如:西南玉米螟(Diatraea grandiosella)和甘蔗螟(Diatraea saccharalis)。这些秸秆食心虫生活在相似的生境下并且会对玉米植株造成相似的破坏。虽然欧洲玉米螟已经被转Bt基因玉米(玉米植株中可以表达Bt毒素)控制住,但是,现在人们日益关心的是由于大面积种子转Bt玉米,欧洲玉米螟有可能对Bt的产生抗性(Gould, 1998;Wolfenbarger and Phifer, 2000)。事实上,在实验室选择条件下,已经有几个欧洲玉米螟群体对Bt毒素产生了抗性(Huang et al., 1997; Bolin et al., 1999)。
Bt毒素的主要靶标是昆虫的中肠,在这里Bt原毒素被中肠道蛋白酶激活转化为具有活性的Bt毒素。激活后的毒素结合到特定的受体上从而表现出毒性(Gill et al., 1992)。这就意味着,昆虫可能通过蛋白酶调节机制和受体调节机制而赋予对Bt毒素的抗性(Oppert et al., 1997; Huang et al., 1999; Li et al., 2004; Lee et al., 1995; Herrero et al., 2001) 。因为Bt毒素和昆虫肠道相互之间的作用是由昆虫肠道内的大量基因产物所决定的,这些基因产物包括许多涉及到与Bt原毒素激活、毒素与受体的结合以及毒素降解相关的蛋白质/酶。这些体系中的任何的改变都有可能影响到Bt的某一特性和效能,可能导致昆虫对Bt的抗性。
我们的研究目标是基于欧洲玉米螟幼虫构建一种肠道特异性EST数据库以及探究有可能与昆虫和Bt相互作用以及Bt抗性有关联的候选基因。本文中,我们列出了对15 000个从欧洲玉米螟幼虫肠道里得到的ESTs的分析和注释。我们讨论了这些ESTs的假定特性,可能存在的生物学功能和分子功能,以及目前我们的ESTs其他昆虫序列的比对分析。这项研究为发展一种可以用来研究昆虫和Bt的相互作用以及欧洲玉米螟Bt抗性的遗传基础的欧洲玉米螟的肠道特异性芯片提供了前体条件。而且,我们得到了52种可能与Bt抗性和毒性的候选基因。从中选出41个通过RT-PCR检测,与欧洲玉米螟的Cry1Ab敏感虫系相比较,我们发现抗性虫系的5个基因表达量下降,10个基因表达增加。这些基因在欧洲玉米螟的Cry1Ab抗性虫系中表达差异可能说明了这些基因与对Cry1Ab产生抗性可能有关联,因此,这也给我们提供了欧洲玉米螟中对Cry1Ab产生抗性机制的一种新的认知。该研究可以作为研究Bt的抗性机制以及研制用于所有密切相关的玉米秸秆食心虫的生物杀虫剂一种模式。
1结果和讨论
1.1欧洲玉米螟的肠道ESTs的构建和分析
首先,我们用从欧洲玉米螟五龄幼虫的整个肠道中纯化得到全部的RNA,利用pPCR-XL-TOPO质粒载体来构建一个cDNA文库。当我们对1 152个cDNA克隆子进行了测序后,我们发现cDNA插入到载体上的片段不够长(平均长度: 441 bp),因此,我们从欧洲玉米螟五龄幼虫的肠道纯化得到的mRNA,用λ Uni-ZAP RX载体构建第二个cDNA文库。这个文库可以为我们提供更长一些的cDNA插入片段(平均长度: 674 bp)。由于λ 文库中的ESTs质量有了重要改良,我们决定使用λ文库作为进一步的ESTs的测序之用。从两个文库中随机挑取了15 000个cDNA克隆子进行测序,仅仅只有不到8%来自质粒文库,而超过92%来自λ文库表1)。
![]()
表1 15 000个欧洲玉米螟幼虫肠道EST分析
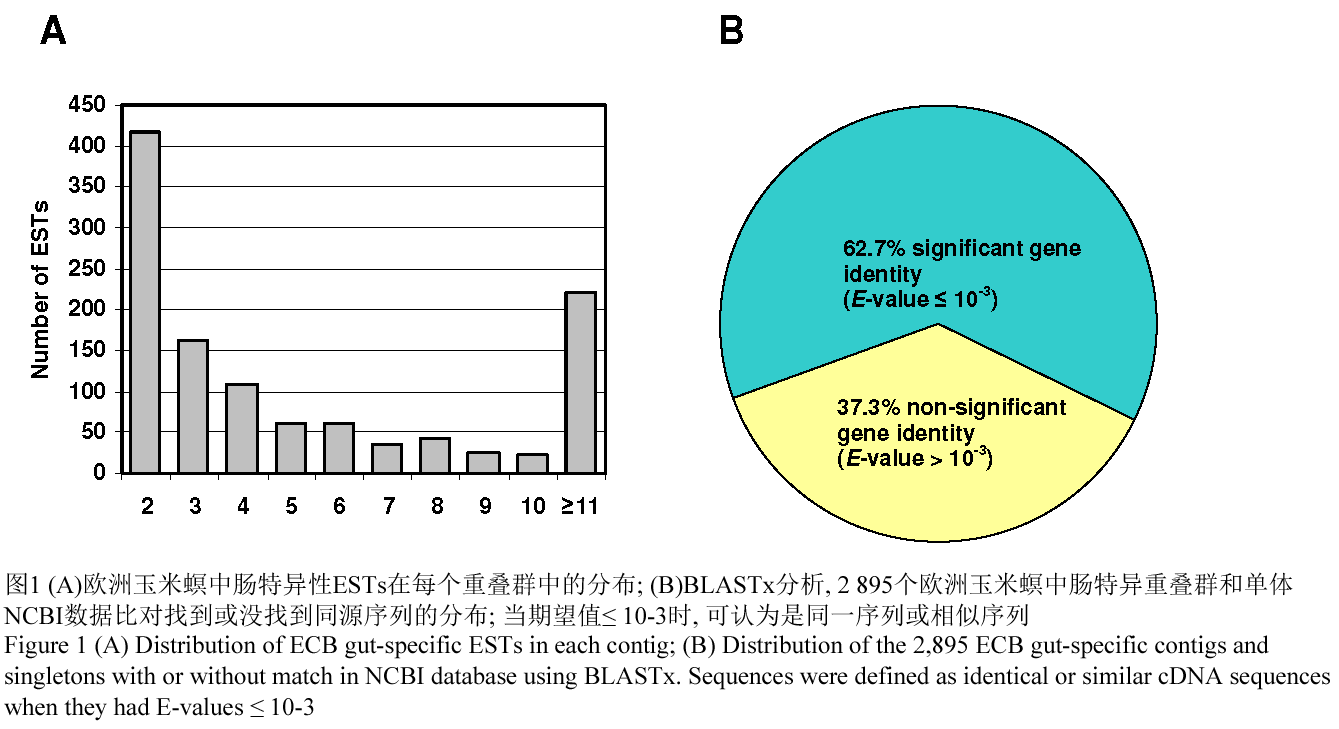
我们对15 000个序列进行分,得到13 066个可读序列(即87.1%的成功率)。这些序列首先去掉载体序列,然后过滤掉长度小于100 bp的序列。用Repeat-Masker and Organelle Masker程序(Masoudi-Nejad et al., 2006)进一步分析,我们除去了另外547个序列。这样,我们总共获得高质量的序列片段有12 519个(占总序列数的83.4%),它们平均长度为656 bp (表1)。这些高质量的序列已经提交到美国国家生物技术信息中心(NCBI)的EST数据库(dbEST)中,GenBank登录号从GH987145到GH999663。使用Sequencher软件(美国密歇根州安拉伯基因编码公司)对这些高质量序列进行冗余及拼接分析,得到2 895个特异的EST序列,其中有1 157个含有2个或者更多个序列的连续序列(重叠群),还有1 738由单一序列组成的单体。大多数的重叠群是由10个或是更少一些的ESTs组合在一起的(图1A)。但是,由于一些高冗余的ESTs的存在,平均起来每一个重叠群是由10.1个EST序列组成的。我们通过用BLASTx分析,从GenBank中非冗余数据库中搜索来定义这些单一序列的假定身份。在2895个特异性序列中,1 816 (62.7%)个序列能找到与他们相似性高的序列,同源期望值≤ 10-3,然而剩余的1 077个序列(37.3%)却没能找到与他们高度相似性的序列(图1B)。
|
|
1.2转录本丰度
生物体内的一种特定基因的转录本丰度可以通过cDNA文库中相应的EST丰度估算出来(Papanicolaou et al., 2005) 。在我们cDNA文库中,最大丰度的ESTs编码胰蛋白酶和糜蛋白酶(表2)。由于这个cDNA文库是由欧洲玉米螟的肠道构建出来的,可以预料其中含有大量的消化酶转录本。丰度最大的重叠群是#0038,它由525个ESTs组成,占整个序列的4%。这个重叠群与胰蛋白酶以前在欧洲玉米螟中与T25前体具有最多相似序列(Li et al., 2005)。其他转录本丰度比较的重叠群有#0026和#0062。#0026重叠群含有197个ESTs并且编码一种假定的糜蛋白酶。#0062重叠群含有137个ESTs,编码一种假定的胰蛋白丝氨酸酶。欧洲玉米螟幼虫肠道中高度表达的基因可能对它们生长和发育有重要影响。然而,由于存在假象的可能,所以我们在对EST的冗余做一般结论的时候必须持谨慎的态度(Kikuchi et al., 2007)。
|
|
.png)
1.3 对开放阅读框(ORF)和假定分泌蛋白的鉴别
用ORF predictor生物软件分析2 895个重叠群和单体,找到ORF的序列。这样做是为了清楚的知道新候选基因的编码容量。在2 895个特有的ESTs中,1 119 (38.7%)个有ORF,大小至少有450 bp。而在这1 119个ORF中,994个假定的蛋白序列(88.8%)与Genbank中的非冗余数据库中已知的蛋白质具有相似性(E value≤10-3)。然而也有125 (11.2%)个假定蛋白质序列在同样的数据库中并没有找到具有明显相似性蛋白序列(图2A)。因此,至少有11.2%的欧洲玉米螟肠道中的编码蛋白质的基因可能是新基因。剩下的ESTs中有1 553个重叠群和单体(53.6%)含有一个小于450 bp的ORF,还有223个(7.7%)并没有ORF。在含有小于450 bp ORF的ESTs中,452 (29.2%)个在非冗余蛋白质数据库中能知道与之相同源性的蛋白质序列,然而,1 101 (70.8%)个找不到相同源性的蛋白质序列。许多序列之所以没有≥450 bp的开放阅读框是因为它们本身序列太短了(大约有650个序列的长度小于450 bp)。其他缺少ORF的序列可能是由于移码突变,cDNA的5′端被切断以及ESTs本身不是来自于mRNA (Whitfield et al., 2002)等原因造成的。
|
|
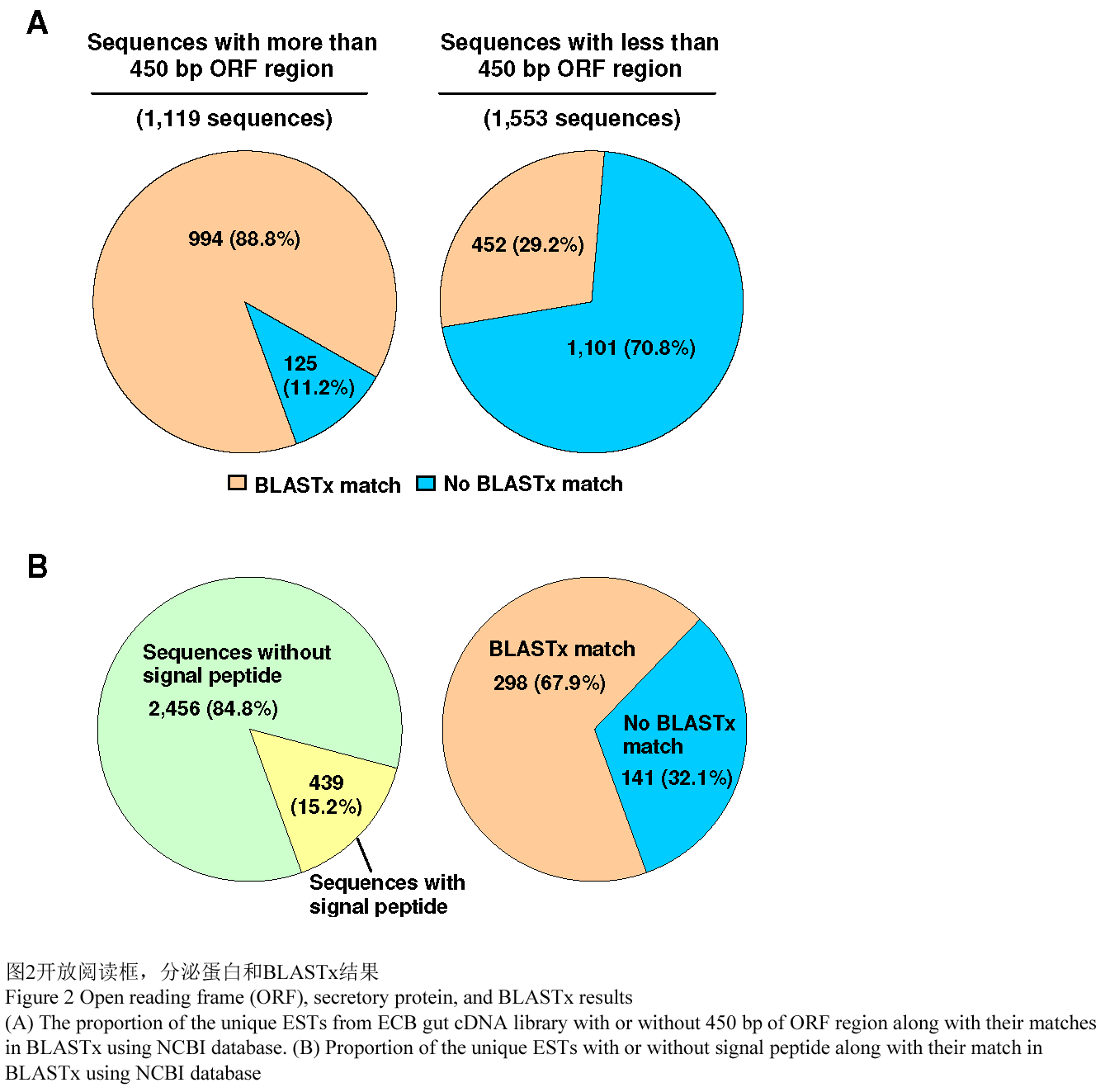
为了鉴定分泌性蛋白质,我们利用SignalP软件(Bendtsen et al., 2004) 对假定蛋白序列进行了检测,鉴定潜在的分泌信号肽。总共有439 (15.2%)个假定基因被预测含有信号肽(图2B)。在假定的分泌蛋白质中,298 (67.9%)个可以从非冗余蛋白质数据库中找到具有同源性的已知蛋白质序列,141 (32.1%)个假定分泌蛋白具有特异性,和任何已知蛋白质都没有显著同源性。这个信息是很有价值的,因为分泌蛋白是在肠道生物过程的一个很重要组成部分(O'Donnell and Blackman, 2005; Adams et al., 1993)。
1.4 欧洲玉米螟肠道ESTs的比对分析
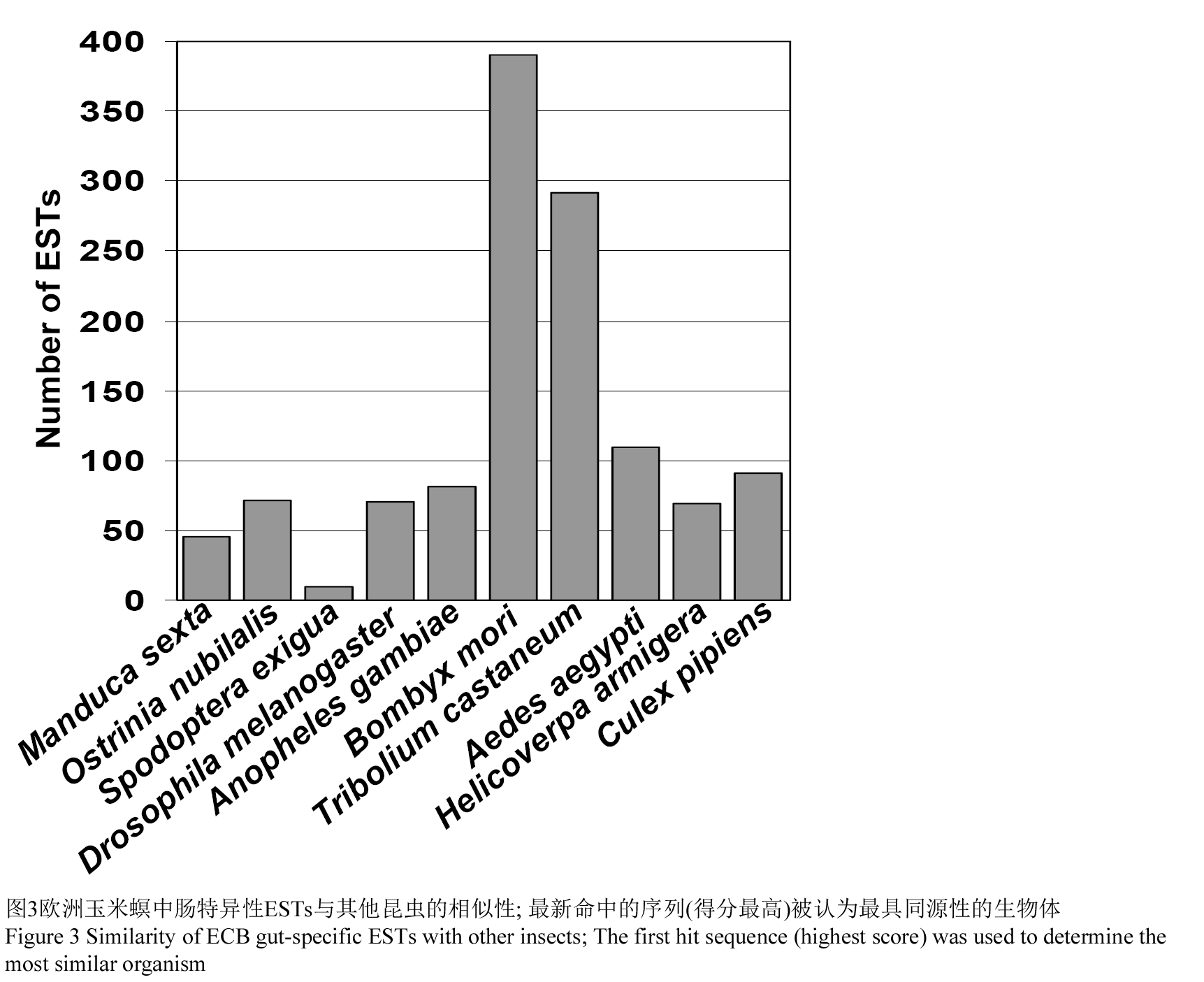
构建EST数据库被认为是一种快速取样生物转录物组的方法,它与全基因组的测序相辅相成(Tosini et al., 2006)。实际上,已经从其他模式生物体内获得许多ESTs。从欧洲玉米螟的幼虫肠道中获得的2 895个重叠群和单体和从其他生物体中得到的序列进行了比对分析。在非冗余数据库中最新命中的序列(得分最高)被认为是同源性最高的生物体。首先命中的序列的最多次的序列来自蚕(390; 13.5%) (图3)。可以解释这样一个事实的是因为桑蚕的基因组已经测序而且有一部分已经注释,并且欧洲玉米螟和桑蚕都是属于鳞翅目的昆虫。首先命中的序列中,数量排在第二位的是赤拟谷盗(290; 10.0%),其后分别是埃及伊蚊(109; 3.8%),尖音库蚊(91; 3.1%)以及冈比亚按蚊(81; 3.8%)。仅仅只2.5%的序列(72)是和预测的欧洲玉米螟的蛋白序列相似性最高。原因很简单,是因为目前欧洲玉米螟在NCBI的数据库中可以用于比对的序列数目很少。
|
|
为了使我们的欧洲玉米螟肠道ESTs能和NCBI数据库中可用的1 745欧洲玉米螟ESTs进行比对,我们使用了BLASTN分析。在我们2 895个重叠群和单单体中,在NCBI数据库中进行BLASTN分析,1 279 (44.2%)能找到同源性高的序列E value≤ 10-3,同时有1 616 (55.8)个是没有找到显著相似性序列。我们的欧洲玉米螟ESTs和NCBI中的已有的欧洲玉米螟dbESTs数据库进行比对分析。我们发现有475个序列(16.4%)的显著的同源性,同源期望值小于E-150 (图4A)。在这里面,88个ESTs (3.0%)同源性期望值低于E-150,23 (0.8%)个同源性期望值在E-100和E-150之间,131 (4.5%)个的同源性期望值是E-100和E-50,152 (5.2%)个的同源性期望值是E-20和E-50还有81 (2.7%)个的同源性期望值在E-5至E-20之间(图4B)。总共有2 420 (83.6%)个ESTs与NCBI数据库中现有的中肠ESTs比对分析,没有找到具有同源性的序列。
|
|
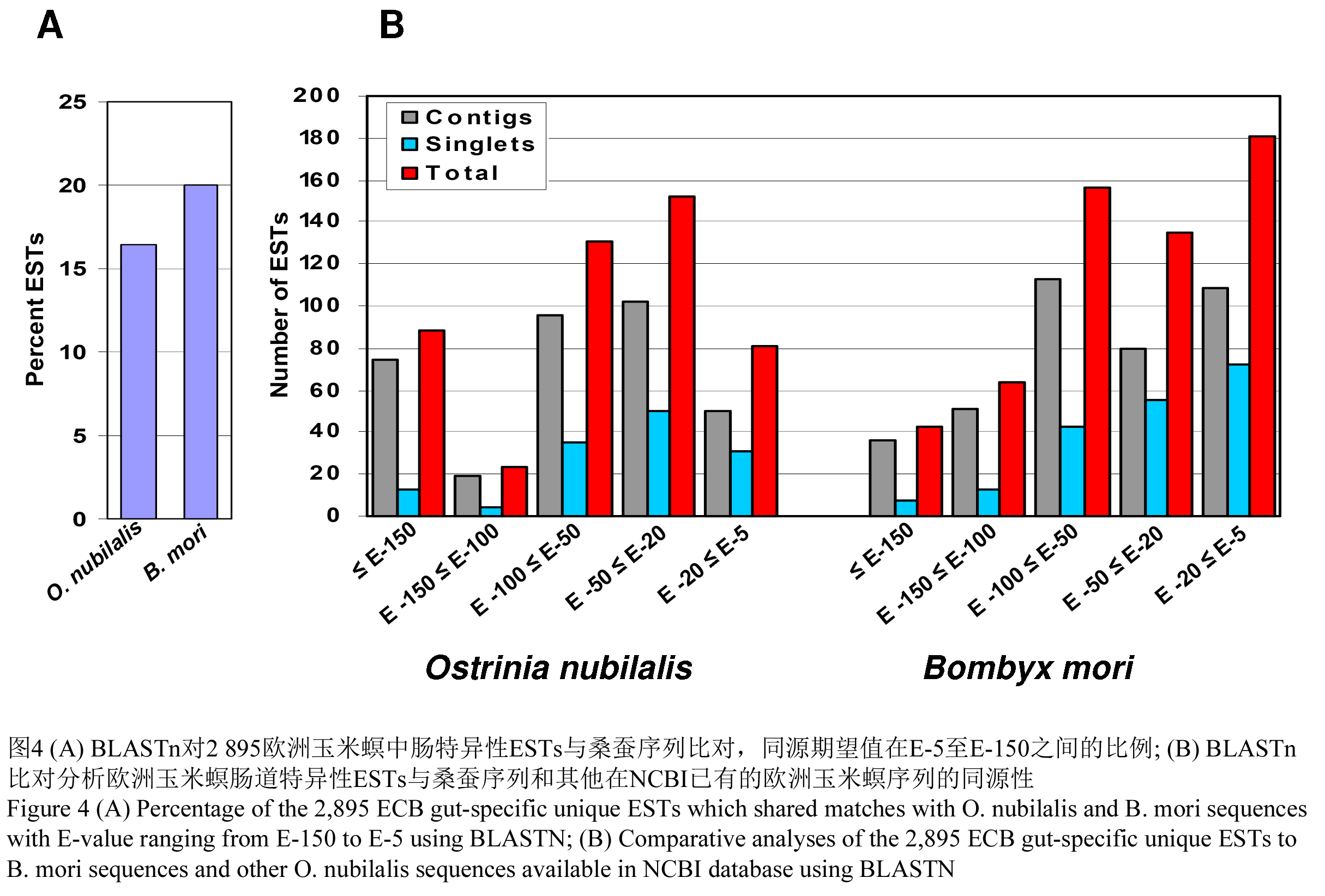
因为桑蚕的基因组没有全部得以注释,我们也对我们的ESTs和所有的桑蚕的ESTs进行了BLASTN比对分析。在2 895个重叠群和单体中,579 (20.0%)个与桑蚕的序列同源性期望值< 10-3 (图4A)。剩余的2 316个ESTs (80.0%)和桑蚕的序列并没有表现出显著的同源性。在579个已经放入数据库的特异ESTs中,43 (7.4%)同源性期望值小于E-150,64 (11.1%)个同源性期望值在E-150和E-100之间,156 (26.9%)个同源性期望值在E-50和E-100之间,135 (23.3%)个同源性期望值在E-20和E-50之间,还有181个序列(31.3%)有同源性期望值在E-5和E-20之间(图4B)。
1.5 基因本体论
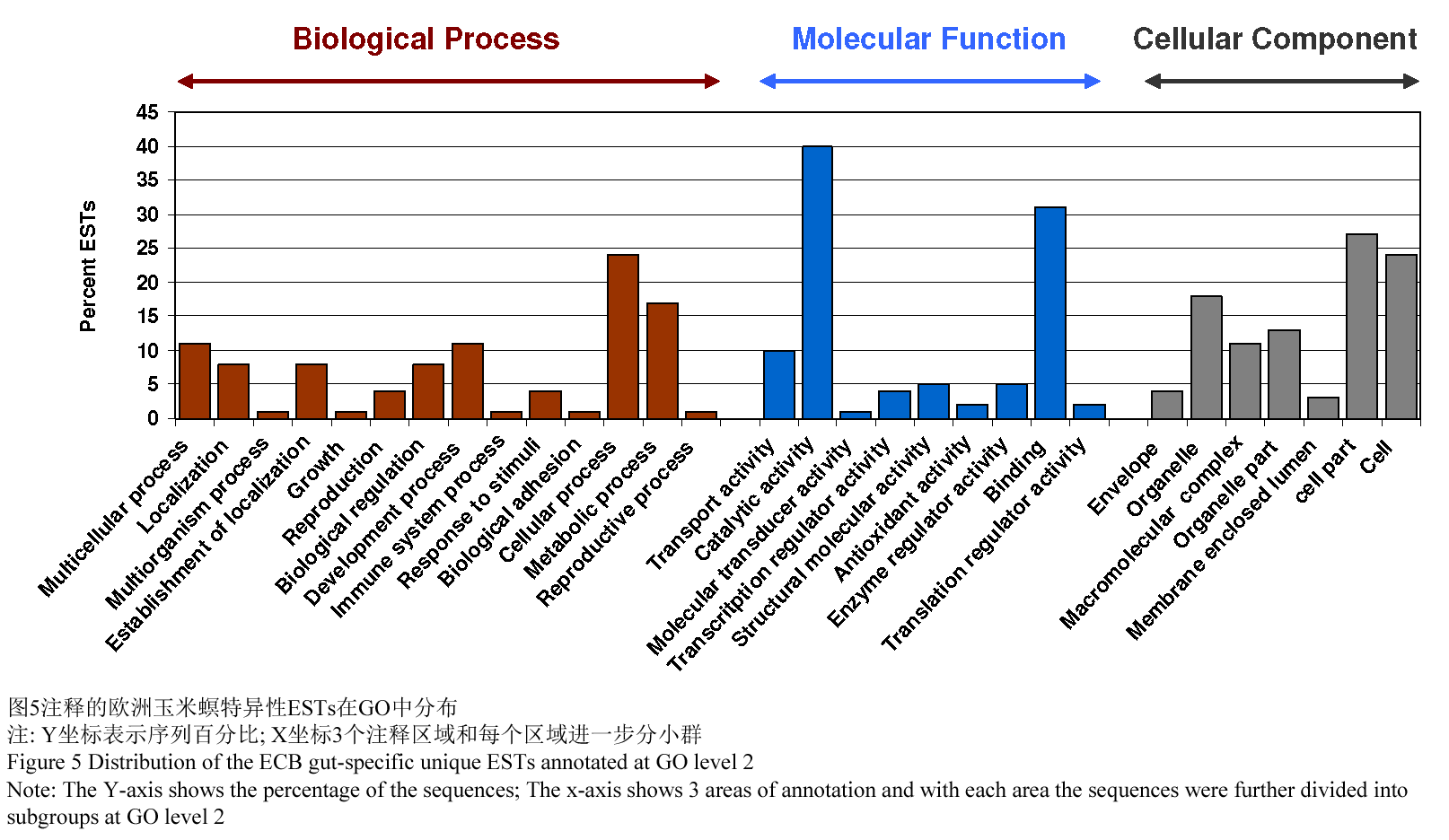
BLAST 2GO 软件是用来获得基因本体论,基因本体论是通过基因本体公司对特异性序列进行比较的专门术语(Ashburner et al., 2000)。在2 895个重叠群和单体中,1 815的比对同源性期望值≤10-3,其中的1 119个ESTs已经绘制了图谱。120个已经绘制了图谱的ESTs同时有基因本体论和酶学委员会的编号。图5表明了EST的生物功能类别,而这些欧洲玉米螟的特异性ESTs都分配到假定的生物过程,分子功能和细胞成分。在生物过程类别中,24.0%是属于细胞进程,接下来17.0%的属于新陈代谢过程,11.0%的生长发育过程,11.0%的多细胞进程以及8.0%属于每一个的生物控制和定位。在分子功能分类中,最大的基因本体论术语(40.0%)是被包括在催化活性里面的,接下来锚定(31.0%),运输活性(10.0%)以及每一个5.0%的酶力控制活性和(9.0%)结构分子活性。在细胞成分的分类中,细胞局部,细胞以及细胞器分别有27.0%,24.0%和18.0%的GO术语。紧跟着是细胞器部分(13.0%),复杂高分子(11.0%),膜结构(4.0%)以及细胞膜封闭腔(3.0%)。
|
|
1.6 与Bt的毒性和抗性有潜在关联的ESTs鉴定
Bt在昆虫中的作用方式主要包括以下几个步骤:摄入Bt原毒素,在肠道内溶解Bt原毒素,原毒素水解激活,毒素与Bt受体结合,细胞隔膜融合,形成微孔,细胞溶解,昆虫的死亡(48)。通过这种作用方式,一个靶标昆虫有可能凭借对Bt与受体相互作用途径进行一个或多个改变来产生对Bt原毒素或毒素的抗性。实际上,关于Bt抗性机制有2种最为普遍观点是蛋白酶调节和受体调节(Ferré and Van, 2002)。我们对从欧洲玉米螟的幼虫肠道中得到的ESTs的分析表明有许多基因与Bt的毒性和抗性有关联(表3)。具体来说,我们鉴定出18假定编码胰蛋白酶ESTs和18个假定编码糜蛋白酶ESTs,它们比对同源性期望值范围分别为2e-16至3e-137和3e-27至3e-149。消化酶水解活性的改变,会影响晶体的溶解或原毒素的激活,从而改变Bt原毒素和毒素的毒性。也降低了毒素的活性(Oppert et al., 1997; Milne and Kaplan, 1993; Oppert et al., 1994; Oppert et al., 1996; Martínez-Ramírez et al., 1996; Keller et al., 1996; Forcada et al., 1996; Forcada et al., 1999)。我们实验室之前的一个研究表明欧洲玉米螟的苏云金芽孢杆菌-抗性虫系对Bt的抗性主要表现与胰蛋白酶活性降低有关(Li et al., 2004; 2005)。这些胰蛋白酶在我们的EST的分析中也有叙述。因此,我们分析欧洲玉米螟幼虫肠道的ESTs展示了更多候选基因,这些候选基因值得进一步分析它们在Bt毒性和抗性中扮演什么样的角色。
|
|
.png)
我们的EST分析显示有13种假定编码氨肽酶ESTs(比对同源性期望值在1e-64至1e-116之间),1个假定编码钙粘着蛋白(比对同源性期望值为1e-35)还有2假定编码碱性磷酸酶(比对同源性期望值在1e-115至1e-131之间)。氨肽酶N,钙粘着蛋白以及碱性磷酸酶已经在其他的昆虫体内发现为Bt的毒素蛋白的结合受体(Herrero et al., 2005; Hara et al., 2003; Jurat-Fuentes and Adang, 2004)。为了验证氨肽酶N在斜纹夜蛾属中具有作为Cry1Ac毒素蛋白受体的功能,我们用RNA干扰技术来减少氨肽酶N的表达。导致昆虫对Cry1Ac毒素敏感性大大降低(Rajagopal et al., 2002)。Gahan等人(Gahan et al., 2001)提出在一种烟芽夜蛾的抗性虫系(YHD2)中,钙粘着蛋白超基因家族被一种反转录转座子介导的插入而中断,而导致该虫系对Bt的Cry1Ac毒素具有高抗性。Fernandez等(2006)人也报道过GPI(糖基磷脂酰肌醇)锚定TLP(碱性磷酸酶)是一种非常重要的受体分子,它涉及到Cry11Aa和埃及伊蚊幼虫中肠细胞的相互作用和毒性。这些研究证明了氨肽酶类,钙粘着蛋白以及碱性磷酸酶可以作为与Bt毒素的受体而与Bt毒性和抗性有关联。因此,本研究对这些Bt受体候选基因的鉴定,让我们可以进一步研究受体介导抗性是否与欧洲玉米螟中对Bt抗性有关联。
1.7欧洲玉米螟对Cry1Ab抗性和敏感虫系的表达谱比较
我们用RT-PCR的方法在欧洲玉米螟的Cry1Ab抗性和敏感虫系中与Bt毒性和抗性相关联的候选基因的表达图进行比较。在52个候选基因中挑选出来的41个基因进行比较,其中15个为假定编码胰蛋白丝氨酸酶,13个编码糜蛋白丝氨酸酶,10个编码氨肽酶,2个编码碱性磷酸酶,还有1个编码钙粘蛋白质,我们发现与敏感性虫系相比,在抗性虫系中有2个胰蛋白酶基因,2个糜蛋白酶基因和1个氨肽酶有明显的表达量减少(图6)。在这些基因中,2个胰蛋白酶基因(重叠群[0907]和欧洲玉米螟-30-CO8)实际上是在抗性虫系中已经丢失了。与之相反,我们发现在抗性虫系中,有3个胰蛋白酶基因和3个糜蛋白酶的基因,2个氨肽酶基因以及2个碱性磷酸酶基因表达量明显增加。增加最显著的有1个胰蛋白酶(重叠群[3395])基因,3个糜蛋白酶(ECB-V-25, 重叠群[0379]以及ECB-23_F02)基因,1个碱性磷酸酶(重叠群[5091]和1个氨肽酶(ECB-D12)基因。
|
|

虽然RT-PCR不是定量分析,但是在欧洲玉米螟的Cry1Ab敏感和抗性虫系中的表达谱中多次出现表达量差异的结果,这就有可能说明它们表达的蛋白对Cry1Ab抗性和欧洲玉米螟敏感虫系和抗性虫系遗传差异方面扮演角色。事实上,已经有人发现一些胰蛋白酶和糜蛋白酶能够激活或抑制Bt的原毒素和毒素,而一些氨肽酶,钙粘着蛋白以及碱性磷酸酶已经被证明是在其他昆虫中扮演着Bt的蛋白质受体的角色。因此,我们的实验结果可能帮助我们坚信候选基因与Cry1Ab抗性有关联,同时也给我们提供了欧洲玉米螟对Cry1Ab抗性机制上的一种新的视角。不过,需要进一步实验来证实它们在欧洲玉米螟对Cry1Ab抗性方面的相关性以及对它们扮演的角色进行解释。
2结论
我们的研究最终构建一个特异性肠道EST的数据库,而这个数据库包括了12 519个高质量的ESTs,这些高质量的ESTs是通过测序一种农业上重要的鳞翅目害虫的15 000个ESTs获得的。就我们所知,这个数据库是鳞翅目昆虫中最大的特异性肠道EST数据库。我们ORF predictor分析表明在我们的数据库中大约11.2%的蛋白质编码基因可能是欧洲玉米螟所特有的,因为这些ORF片断至少有450 bp而它们和NCBI数据库中已知序列没有显著同源性。我们也鉴定了52个与Bt的毒性和抗性有关联的候选基因。这些基因编码胰蛋白酶,与糜蛋白酶,氨肽酶,钙粘蛋白以及碱性磷酸酶。此外,我们用RT-PCR方法来检测比对对cry1Ab抗性虫系中的41个具有代表性的候选基因表达谱,15个候选基因具有明显的表达差异,在抗性虫系中,有5个表现为明显表达量减少,10个表现为明显表达量增加。这个结果可能会帮助我们进一步缩小与Cry1Ab抗性有关联的候选基因的范围,并且给我们提供一个对欧洲玉米螟的Bt抗性机制的新的视角。
我们现在正在尝试用我们的特异性ESTs以及在GenBank中已有的欧洲玉米螟的肠道特异性序列来研发一种基因芯片。这种基因芯片技术将可以帮助我们分析基因表达量的全局变化对Bt的原毒素和毒素的反应。它将使我们能对欧洲玉米螟的Bt抗性和敏感虫系的遗传差异进行分析。我们关于欧洲玉米螟的基因组信息可以作为一种宝贵的资源用于鉴定欧洲玉米螟肠道中关键或敏感的基因,可能产生有用新毒素的生理性靶标用于害虫的防治。
3方法
3.1昆虫饲养及解剖
使用欧洲玉米螟的Bt敏感虫系KS-SC 构建EST库。这个虫系是1995年,从St. John, Kansas附近的玉米田里采集到的卵块而获得的。参考黄等人的方法(Huang et al., 1997),在堪萨斯州立大学的实验室内,用人工饲料将这个采集虫系一直饲养至今。欧洲玉米螟的抗性虫系是2001年从Kandiyohi Co., MN非Bt杂交材料田里采集获得的126个滞育幼虫。抗性虫系最初来源14只幼虫,这14只幼虫是在暴露于用于鉴定对Cry1Ab敏感性潜在变化实验的cry1Ab毒素浓度下存活的下了的(Siegfried et al., 2007; Crespo et al., 2009)。为了使近亲繁殖和奠基者效应降到最低,将抗性虫系和同一批收集的敏感虫系回交2次。因为抗性是不完全隐形遗传并且涉及到多种因素(Crespo et al., 2009) 。F1代随机交配得到具有抗性因子重组子的F2,从中可以筛选到抗性基因型。这些虫子在cry1Ab浓度相当于F1代半致死量浓度2~3倍下进行选择性筛选(Marçon et al., 1999) 。筛选试验是将所有敏感的纯合子和大部分杂合子去除掉。筛选试验中存活下来的虫子将进行第二轮的回交、随机交配和筛选。6个世代以后,使用的Cry1Ab的浓度逐渐增加,F10代时,cry1Ab的使用浓度达到750 ng/cm2,这个浓度实际上可以杀死全部的F1代幼虫了。F17时,重复筛选得到虫具有的抗性已经超过F1 800倍了。解剖5年龄幼虫,将肠道用DEPC处理,然后用蒸馏水清洗,放入TRI reagent™ (Molecular Research, Inc., Cincinnati, OH),存放在-80°C条件下备用。
3.2 cDNA文库的构建和测序
使用TRI试剂将全部的RNA从欧洲玉米螟幼虫的整个肠道中提取出来。质粒文库是使用Creator SMART™ cDNA文库试剂盒(Palo Alto, CA)构建,我们按照产品的说明书进行构建,只进行一处的修改,那就是我们用TOPO TA克隆试剂盒(Invitrogen, Carlsbad, CA)直接将PCR产物插入到pPCR-XL-TOPO质粒载体中取代原先使用的噬菌体载体。λ-文库是利用ZAP-cDNA合成试剂盒以及ZAP-cDNA Gigapack III gold 克隆试剂盒(Stratagene, La Jolla, CA),按照产品说明书的来构建。简言之,使用poly(A) RNA合成双链cDNA。使用CL-2B琼脂糖凝胶层析柱分离不同大小的cDNA,然后连接到λ Uni-ZAP XR载体上。重组的DNA用Gigapack III gold抽提物来包装,并且构建文库涂布在LB/琼脂平板上。插入到λ Uni-ZAP XR载体重组质粒采用ExAssist辅助噬菌体进行体内分离并传递pBluescript SK噬菌体载体进行亚克隆。为了测序克隆子,5'和3'的测序分别使用引物M13R和引物M13F。质粒DNA的分离是使用Qiagen Bio Robot 3000,测序使用了ABI3700 DNA analyzer。
3.3 EST分析和注释
使用在线软件EGassembler (Masoudi-Nejad et al., 2006)对DNA序列进行了预加工。具体来说,序列清洗程序就是用来剪切掉EST上的载体和衔接子的序列。Repeatmasker程序是利用Drosophila Repbase repeat来修饰穿插重复序列和低复杂度序列区域。序列进一步修饰,通过与与NCBI载体数据库比对进行载体修饰,通过与线粒体数据库比对进行细胞器修饰。经过预加工的EST序列通过Sequencher软件(Gene Codes Corp., Ann Arbor, MI)进行装配。装配好的的ESTs的ORF区域通过ORF predictor软件(Min et al., 2005) 被鉴别出来,用SignalP软件(Bendtsen et al., 2004)寻找信号肽顺序来鉴定分泌蛋白序列。基因本体论的注释的来源于Blast2GO (http://www.blast2go.de/; Conesa et al., 2005)。
3.4 ESTs的对比分析
欧洲玉米螟的特异性ESTs与其他的生物体进行了序列同源性的比对分析。在GenBank数据库中比对得分最高相关的生物被挑选出来。欧洲玉米螟的肠道ESTs也与家蚕的序列进行比对同时采用BLASTN比对与数据库目前已有欧洲玉米螟的序列进行了比对分析,只分析同源性期望值到10-3为止的序列。
3.5 通过RT-PCR获得表达谱
在欧洲玉米螟对Cry1Ab敏感性和抗性的虫系中,从52个候选基因中选出41个通过使用RT-PCR,比较它们中清晰可见的基因表达谱。这些基因的筛选仅仅是基于通过ESTs分析从不同的基因群中选出的代表性。解剖饲养过一天欧洲玉米螟5龄幼虫,从每个虫系获得(Cry1Ab敏感以及抗性虫系)获得4份中肠,然后用TRI试剂盒(Sigma, St. Louis, MO)处理获得总RNA,再用TURBO™DNA聚合酶(Ambion, Austin, TX)来处理,去除基因组DNA的污染。我们取出3 mg的RNA利用SuperScript® IIIFirst-Strand合成体系(Invitrogen, Carlsbad, CA)来合成cDNA的第一条链。从总RNA中制备的cDNA是用来作RT-PCR反应的模板的。生物学复制最小量的2个片段被用来作为所有PCR反应的引物对。所有的胰蛋白酶(除了欧洲玉米螟-30_C08)糜蛋白丝氨酸酶,碱性磷酸酶和RPS3基因进行25个PCR反应循环,而氨肽酶以及钙粘蛋白基因要进行27个PCR反应循环。一种胰蛋白丝氨酸酶(ECB-30_C08)需要33个PCR循环,因为这个基因如果PCR反应循环太少,就不能在琼脂糖凝胶板清晰的显现出来。每个PCR反应要按照上面所述的循环次数来进行,每一个反应条件为:94°C变性30 s,55°C退火60 s以及72°C延伸60 s。PCR的正向反应引物序列和41个候选基因中的每一个的基因的PCR产物预计片度大小都将附假材料中列出。
作者贡献
CK指导了本研究的主要部分包括:实验设计,cDNA文库的构建,EST分析,RT-PCR的分析以及初稿的准备工作。YCZ参与了实验的设计,EST的测序和EST数据的前期分析工作。MSC促进项目的开展,在EST测序上有稳定合作关系并未初稿的准备上提供了帮助。LLB参加了实验设计,昆虫饲养以及初稿的准备等工作。RAH参与了这个项目的发展工作以及实验设计。JY协助EST的测序以及分析。BDS和ALBC提供了材料以及参与了数据库的分析和初稿的准备。SM参与实验设计以及初稿的准备。KYZ负责协调整个项目并且参与了实验设计,EST的分析以及初稿的准